
-
Mail us:
editor@raftpubs.org
Indexing & Abstracting
Full Text
Research ArticleDOI Number : 10.36811/ijbm.2019.110006Article Views : 2153Article Downloads : 18
Chromosomal Q-heterochromatin Variability and Sport
Ibraimov AI
Laboratory of Human Genetics, National Center of Cardiology and Internal Medicine, Bishkek, Kyrgyzstan
*Corresponding author: Ibraimov AI, Laboratory of Human Genetics, National Center of Cardiology and Internal Medicine, Bishkek, Kyrgyzstan. Email: ibraimov_abyt@mail.ru
Article Information
Aritcle Type: Research Article
Citation: Ibraimov AI. 2019. Chromosomal Q-heterochromatin Polymorphism and Sport. Int J Biol Med. 1: 45-52.
Copyright: This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Copyright © 2019; Ibraimov AI
Publication history:
Received date: 13 March, 2019Accepted date: 27 March, 2019
Published date: 29 March, 2019
Abstract
In the process of studying the variability of chromosomal Q-heterochromatin regions (Q-HRs) in human populations living permanently in different climatogeographic conditions of Eurasia and Africa, as well as in newcomers, who have successfully adapted to some extreme natural conditions, the data were obtained that, perhaps, the amount of this genetic material in the genome may be important for some sports. In particular, it turned out that in the genome of mountaineers the number of chromosomal Q-HRs was significantly lower than in the control sample and was close to that of the high-altitude natives. The same results were obtained in a comparative study of newcomers who have been working in the extreme climatic conditions of the Far North of West Siberia. We believe that these observations can be explained by cell thermoregulation (??) the level of which depends on the number of chromosomal Q-HRs. The essence of ?? is elimination of the temperature difference between the nucleus and cytoplasm. The effect of CT can be indirectly assessed by the level of the body heat conductivity (BHC). An individual with high BHC cannot make much progress in mountaineering and water sports due to the fact that their body cools rapidly. However, this individual can be more successful in sports which require effective heat-loss.
Keywords: Chromosomal Q-heterochromatin; Cell thermoregulation; Body heat conductivity; Sport genetics; Marathon, Mountaineering
Introduction
The study of the literature on the possible role of heredity in sports shows that there are no specific studies on this problem, that is, there are no works that would show the relationship between a certain genotype and sport (phenotype). However, the absence of such studies does not mean that heredity plays no role in sports. Perhaps here we still know more about the role of inborn features of the athlete than his heredity.
In the process of studying the variability of chromosomal Q-heterochromatin regions (Q-HRs) in human populations living permanently in different climatogeographic conditions of Eurasia and Africa, as well as in newcomers, who have successfully adapted to some extreme natural conditions (high altitudes and the Far North of Eastern Siberia), the data were obtained that, perhaps, the amount of this genetic material in the genome may be important for some sports. In particular, it turned out that in the genome of mountaineers the number of chromosomal Q-HRs was significantly lower than in the control sample and was close to that of the natives of high-altitude Pamir and Tien-Shan [1]. The same results were obtained in a comparative study of newcomers (oil-borers working in the open air in the oil and gas fields of the Yamal Peninsula of Eastern Siberia) who have been working more than five years in the harsh climatic conditions of the Far North, the indigenous inhabitants of Siberia and their children [2]. A fundamental feature of chromosomes of higher eukaryotes, including man, is the presence of two evolutionary consolidated types of genetic material: euchromatin and heterochromatin. Euchromatin - the conservative portion of the genome - contains transcribed structural genes, while heterochromatin - the variable portion of the genome - predominantly consists of nontranscribed repeated DNA sequences. About 15%-20% of the human genome is composed of heterochromatin regions [3]. To date, two types of heterochromatin are known: C- and Q-heterochromatin [4]. There are several significant differences between them: 1) C-heterochromatin regions (C-HRs) is found in the chromosomes of all the higher eukaryotes, while Q-heterochromatin regions (Q-HRs) - only in man (Homo sapiens), the chimpanzee (Pan troglodytes) and gorilla (Gorilla gorilla); 2) C-HRs are known to be invariably present in all the chromosomes of man, varying mainly in size and location; 3) despite the fact that chromosomal Q-HRs exist in the genome of three higher primates, their broad quantitative variability is only inherent in human populations; 4) individuals in population differ in the number of chromosomal Q-HRs in the genome; 5) chromosomal Q-HRs does not change during ontogenesis and show normal Mendelian inheritance[1-3,5-20].
Despite the fact that in human karyotype there are 13 loci in which Q-HRs can be detected, i.e., there could theoretically exist individuals with 25 Q-HRs in their genome, but such cases have not as yet been reported. In individuals of a population the number of Q-HRs on the autosomes usually ranges from zero to ten [11,12,16]. The distribution of Q-HRs variability of chromosomes in populations is usually described in the form of two main quantitative characteristics: the distribution of Q-HRs in a population, i.e., distribution of individuals having different numbers of Q-HRs in the karyotype regardless of the location and the mean number of Q-HRs per individual. These differences proved to be related to features of the climatic-and-geographic conditions of the place of permanent residence, and not to racial and ethnic composition of the population.
We examined a large sample (n=385) of athletes representing the national team of mountaineers, many of whom participated in the conquest of Everest [1]. The samples of mountaineers studied during various seasons were very different in nationality, sex and age. They were compared as to the distribution and the mean number of chromosomal Q-HRs variability per individual. No significant differences were observed between the athletes as regards all quantitative characteristics of Q-HRs variability. Among mountaineers’ athletes of the Russian nationality made the majority. This sample of mountaineers we used to compare with the control. As can be seen from Table 1 there are statistically significant differences between Russian mountaineers and Russian controls in the distribution and in the mean number of Q-HRs per individual between these samples.
| Table 1: Distribution and the mean number of chromosomal Q-HRs per individual in mountaineers and Russian controls. | ||
|---|---|---|
| Number of Q-HRs | Mountaineers (n=277) | Controls (n=200) |
| 0 | 46 | 9 |
| 1 | 81 | 29 |
| 2 | 100 | 49 |
| 3 | 39 | 54 |
| 4 | 9 | 34 |
| 5 | 2 | 24 |
| 6 | 4 | |
| 7 | 1 | |
| Total Q-HRs | 444 | 572 |
| Mean number of Q-HRs | 1.60 ± 0.06; | 2.86 ± 0.10; |
Of interest is the fact that the mean number of Q-HRs per individual was higher in the high-altitude Kyrgyz sample than in the mountaineers (whereas we initially assumed that in this case we should fail to find any significant differences), also it was lower than in the Russian controls, but in both cases the differences were statistically significant (Table 1,2).
| Table 2: Distribution and the mean number of chromosomal Q-HRs per individual in mountaineers and Kyrgyz (natives of high-altitude Pamir and Tien-Shan) controls. | ||
|---|---|---|
| Number of Q-HRs | Kyrgyz (n = 284) | Mountaineers (n = 385) |
| 0 | 32 | 57 |
| 1 | 67 | 116 |
| 2 | 87 | 141 |
| 3 | 56 | 56 |
| 4 | 30 | 13 |
| 5 | 12 | 2 |
| Total Q-HRs | 589 | 628 |
| Mean number of Q-HRs | 2.07 ± 0.08 | 1.63 ± 0.05 |
The circumpolar zone of Eurasia is inhabited by people of several nationalities which differ significantly from each other in origin, culture and tenor of life. Over the past century this region has been exploited by newcomers, mainly Russians. Two Russian groups were investigated, namely (1) oil-industry borers, who had been permanently engaged in heavy physical work for 5 or more years, and (2) immigrants from various regions of the USSR, together with pupils of schools.
The distribution and the mean number of Q-HRs per individuals were compared in natives. No significant differences were evident in natives of Western Siberia [2]. Therefore, the natives were combined into one group in order to compare it with newly arrived oil-borers who had become adapted to the extreme conditions of the North (Table 3). The last two lines of Table 3 contain information on the resemblances and differences between the samples as to the distribution and the mean number of Q-HRs per individual. There are statistically significant differences between the natives of the northern part of West Siberia, Russian children and Russian controls, in regard to the quantitative characteristics of chromosomal Q-HRs (pairs I, II and I, IV, respectively). There were no such differences between the indigenous people and the oil-borers (pairs I, III). Statistically significant differences were also found between the oil-borers, Russian children and Russian controls (pairs II, III and III, IV) as well as between Russian children and Russian controls (pairs II, IV).
Thus, the number of chromosomal Q-HRs in the genome of oil-borers, who were selected Russians well adapted to the climatic conditions of the northern part of West Siberia, is very similar to that of the Siberian aborigines, but differs significantly from the controls who were Russians from moderate latitudes. Of interest, in our opinion, is also the fact that between Russian children, who, as a rule, are migrants of the first or second generations, and Russian controls there are some differences in the number of Q-HRs in their genome. We are inclined to explain these facts by the possible selective value of Q-heterochromatin in human adaptation to extreme environmental factors of far north of Eurasia.
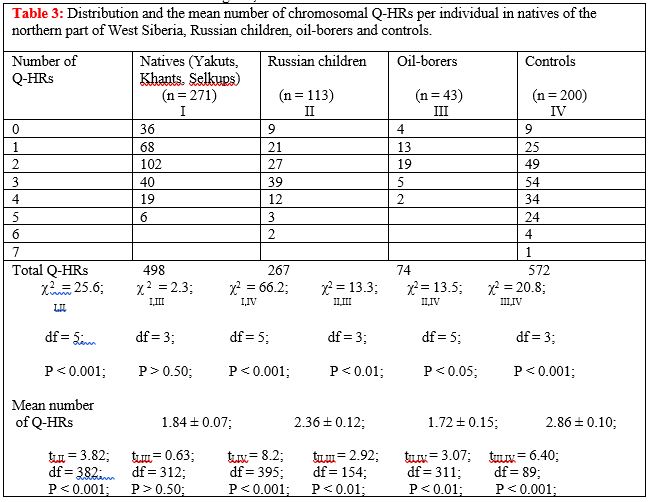
We believe that our studies of the chromosomal Q-HRs variability in the indigenous population of the northern part of West Siberia and in new migrants, albeit indirectly, point to the possible role of the amount of Q-heterochromatin in the human genome in certain sports. Of course, the work of oil-borers is difficult to compare with the physical activity experienced by a professional athlete. But judging by the fact that not all individuals who came to earn good money at the northern part of West Siberia, were able to work more than five years indicates about a serious test of the human body extreme climate of the Far North of Eurasia. It is known that the problems with human adaptation to the extreme climate of the North are primarily associated with the maintenance of temperature homeostasis in the body. This problem occurs when conquering high altitudes by climbers, as well as during marathon races.
There are studies that individual performance depends on endogenous factors such as anthropometric, genetic and physiological characteristics [21,22] and environmental factors that affect all athletes [23-25]. In this regard, it is of interest to observe the athletes who are engaged in marathon running [26]. For example, athletes’ nationalities were categorized into one of the six world regions: Africa, Asia, Europe, North America, South America and Oceania. For each year and both sexes, the contribution of each continent in the top 100 was calculated. It turned out that the best performances by men in marathons were by athletes from East Africa (mainly Kenya and Ethiopia) and this trend has been progressive from 1990 to 2011 (16% to 94%). Conversely, other nationalities, initially dominant in the list of top 100 performers (from Europe, Asia, South America, North America and Oceania), are increasingly less prominent over the same period, 84% in 1990 to 6% in 2011. This change in domination also occurred in the speed of the 10 best performers from each continent since 1990. As for the role of seasonality, it turned out that it has two peaks: the first peak occurs in weeks 14, 15, 16 and 17 (April) while the second occurs during weeks 41, 42, 43 and 44 (October). These eight weeks contain 48.7% of women’s performances (27.6% for April and 21.1% for October) and 54.4% of men’s performance (29.4% for April and 25.0% for October). Performances during weeks 25 to 32 (from early June to late August) represent less than 1% of the total (for more details see [26].
There are other facts indirectly confirming the possible role of heredity in sports. The example, more and more countries situated at southern latitudes have started taking part in the world sport movement lately. The most notable in this process is that, natives of this region achieve great success in sports, requiring (in addition to other factors) effective heat-loss (football, professional boxing and marathon race). While sportsmen from northern latitudes prevail in water and winter sports and also in mountaineering.
Comparative tests for endurance of the “whites” and “blacks” to physical load in conditions of heat and high humidity demonstrate superiority of Negroes even over those “whites” who are used to working in such conditions. On the other hand, the experience of war in Korea showed that frostbite occurs much more often with Negro soldiers than with the “whites”. Long-term experience of the Indian medical officers in the Himalayas shows that the South Indians are physiologically more susceptible as compared to Gorkhas and North Indians under identical environmental conditions and that the high-altitude population is more resistant to cold injures. It was shown that South Indians are more susceptible to frostbite than other ethnic groups of this country [27]. At last, high-altitude pulmonary edema (HAPE) occurs in unacclimatized individuals who are rapidly exposed to altitude in excess of 2500 m above sea level. However, the question remains: why HAPE develops only in some individuals, who are rapidly exposed to high-altitudes? Our experience in studying the genetics of human adaptation to high-altitude shows that the hereditary factor may plays a role.
Discussion
We believe that all of the above observations, directly or indirectly related to sports, can be explained by cell thermoregulation. Based on study of chromosomal HRs the hypothesis of thermoregulation at the cell level has been advanced. The essence of hypothesis of cell thermoregulation (??) is elimination of the temperature difference between the nucleus and cytoplasm when the nucleus temperature becomes higher than the cytoplasm temperature [28-30]. The condensed chromatin (??) localized between a nucleus and cytoplasm is made of different types of chromosomal HRs. For this reason, ?? is subject to wide variability in population. Obviously, the density of the ?? packing depends on the quantity and quality of chromosomal HRs in its structure that can affect upon its heat-conducting ability.
It has been experimentally shown that the effect of CT can be indirectly assessed by the level of the body heat conductivity (BHC). In particular, we were able to show that the amount of chromosomal Q-HRs influence on the level of the human BHC. In other words, there are some parallels in the distribution of the number of chromosomal Q-HRs and variability of BHC at the level of human populations [31].
For example, southerner’s bodies have relatively high heat conductivity [32] it is not surprising, that they are successful in sports, which require effective heat-loss. Indeed, a sportsman with high heat conductivity cannot make much progress in mountaineering and water sports due to the fact that their body cools rapidly. However, this sportsman can be more successful in sports which require effective heat-loss.
Thus, our own research and literature data do not exclude the possible role of heredity in sports. Usually speaking of heredity implies structural genes. However, our data indicate the participation of the non-genetic part of the genome in human adaptation to cold, heat and high-altitude climate. Since the maintenance of temperature homeostasis in the body is critical, there is nothing surprising in the assumption that athletes achieve outstanding success in sports such as mountaineering, professional boxing and marathon running the role of heredity can be very important.
References
- Ibraimov ?I, Kurmanova GU, Ginsburg ??, et al. 1990. Chromosomal Q-heterochromatin regions in native highlanders of Pamir and Tien-Shan and in newcomers. Cytobios. 63: 71-82. [Ref.]
- Ibraimov ?I, Axenrod ?I, Kurmanova GU, et al. 1991. Chromosomal Q-heterochromatin regions in the indigenous population of the Northern part of West Siberia and in new migrants. Cytobios. 67: 95-100. [Ref.]
- Stanyon R, Studer M, Dragone A, et al. 1988. Population cytogenetics of Albanians in Cosenza (Italy): frequency of Q- and C-band variants. Int J Anthropol. 3: 19-29.[Ref.]
- Prokofyeva-Belgovskaya AA. 1986. Heterochromatic Regions of Chromosomes (in Russian). Moscow, Nauka. [Ref.]
- Paris Conference. 1971. supplement (1975). Standartization in human cytogenetics. Birth Defects: Original Article Series, XI, 1-84. The National Foundation, New York.[Ref.]
- Chiarelli B, LinC.C. 1972. Comparison of fluorescence patterns in human and chimpanzee chromosomes. Genet Phaenen. 15: 103-106.[Ref.]
- Pearson PL. 1973. The uniqueness of the human karyotype. In ?. Caspersson, & L. Zech (Eds.), Chromosome identification techniques and application in biology and medicine (p. 145). New York, London. Academic Press. [Ref.]
- McKenzie W?, Lubs ??. 1975. Human Q and ? chromosoma1 variations: distribution and incidence. Cytogenet Cell Genet. 14: 97-115. [Ref.]
- Buckton KE, ORiordan ML, Jacobs PA, et al. 1976. C- and Q-band polymorphisms in the chromosomes of three human populations. Ann Hum Genet. 40: 90-112. [Ref.]
- Lubs HA, Patil SR, Kimberling WJ, et al. 1977. Racial differences in the frequency of Q- and C-chromosomal heteromorphism. Nature. 268: 631-632. [Ref.]
- Al-Nassar ??, Palmer ?G, Connealy P?, et al. 1981. The genetic structure of the Kuwaiti population. II. The distribution of Q-band chromosomal heteromorphisms. ?um Genet. 57: 423-427. [Ref.]
- Ibraimov ?I, Mirrakhimov ??, Nazarenko S?, et al. 1982. ?uman chromosomal polymorphism. I. Chromosomal Q-polymorphism in Mongoloid populations of Central Asia. Hum Genet. 60: 1-7. [Ref.]
- Ibraimov ?I, Mirrakhimov ??. 1982. Human chromosomal polymorphism. III. Chromosomal Q-polymorphism in Mongoloids of Northern Asia. Hum Genet. 62: 252-257. [Ref.]
- Ibraimov ?I, Mirrakhimov ??. 1982. Human chromosomal polymorphism. IV. Q-polymorphism in Russians living in Kirghizia. Hum Genet. 62: 258-260. [Ref.]
- Ibraimov ?I, Mirrakhimov ??. 1982. Human chromosomal polymorphism. V. Chromosomal Q-polymorphism in African populations. Hum Genet. 62: 261-265. [Ref.]
- Ibraimov ?I, Mirrakhimov ??. 1985. Q-band polymorphism in the autosomes and the Y chromosome in human populations. In: “Progress and Topics in Cytogenetics. The Y chromosome. Part ?. Basic characteristics of Y chromosome”. ?. ?. Sandberg (Ed). Alan R. Liss, Inc., New York. USA. 213-287.[Ref.]
- Ibraimov AI, Karagulova GO, Kim EY. 1997. Chromosomal Q-heterochromatin regions in indigenous populations of the Northern India. Ind J Hum Genet. 3: 77-81.[Ref.]
- Ibraimov AI, Akanov AA, Meymanaliev TS, et al. 2013. Chromosomal Q-heterochromatin polymorphisms in 3 ethnic groups (Kazakhs, Russians and Uygurs) of Kazakhstan. Int J Genet. 5: 121-124. [Ref.]
- Kalz L, Kalz-Fuller ?, Hedge S, et al. 2005. Polymorphism of Q-band heterochromatin; qualitative and quantitative analyses of features in 3 ethnic groups (Europeans, Indians, and Turks). Int J Hum Genet. 5: 153-163. [Ref.]
- Decsey K, Bellovits O, Bujdoso G.M. 2006. Human chromosomal polymorphism in a Hungarian sample. Int J Hum Genet. 6: 177-183. [Ref.]
- Lippi G, Favaloro EJ, Guidi GC. 2008. The genetic basis of human athletic performance. Why are psychological components so often overlooked? J Physiol. 586: 3017. 3019-3020. [Ref.]
- Macarthur DG, North KN. 2005. Genes and human elite athletic performance. Hum Genet. 116: 331-339. [Ref.]
- Ely MR, Cheuvront SN, Roberts W O, et al. 2007. Impact of weather on marathon-running performance. Med sci sport exe. 39: 487-493. [Ref.]
- Galloway SD, Maughan RJ. 1997. Effects of ambient temperature on the capacity to perform prolonged cycle exercise in man. Med Sci Sport Exe. 29: 2-12. [Ref.]
- Vihma T. 2010. Effects of weather on the performance of marathon runners. Int J Biometeor. 54: 297-306. [Ref.]
- Marc A, Sedeaud A, Guillaume M, et al. 2013. Marathon progress: demography, morphology and environment. J Sports Sci. 1-9.[Ref.]
- Mathew L. 1992. Severe Cold at High Altitude and Methods of Prevention of its III Effects. In: SK. Machanda, W. Selvamurthy, V. Mohan Kumar (eds): Advances in Physiological Sciences. New Dehli, Macmillan India LTD. 338-345.[Ref.]
- Ibraimov AI. 2003. Condensed chromatin and cell thermoregulation. Complexus. 1: 164-170. [Ref.]
- Ibraimov AI. 2017. Cell Thermoregulation: Problems, Advances and Perspectives. J Mol Biol. Res. 7: 58-79. [Ref.]
- Ibraimov ?I. 2019. Human adaptation: why only genes?. Int J Biol Med. 1: 22-33.[Ref.]
- Ibraimov ?I, Mirrakhimov ??, Axenrod ? I, et al. 1986. Human chromosomal polymorphism. IX. Further data on the possible selective value of chromosomal Q-heterochromatin material. Hum Genet. 73: 151-156. [Ref.]
- Ibraimov AI, Akanov AA, Meimanaliev TS, et al. 2014. Human Chromosomal Q-heterochromatin Polymorphism and Its Relation to Body Heat Conductivity. Int J Genet, 6: 142-148.[Ref.]
- Ibraimov AI, abaldiev SK. 2007. Condensed chromatin, cell thermoregulation and human body heat conductivity. J Hum Ecol. 21: 1-22.[Ref.]
- Yamada ?, Hasegawa ?. 1978. Types and frequencies of Q-variant chromosomes in ? Japanese population. ?um Genet. 44: 89-98. [Ref.]
- Pearson PL. 1977. Pattern of bands, polymorphism and evolution of primates. In J. J. Yunis (Ed.), Molecular structure of human chromosomes. Acad. Press.[Ref.]


















